1. INTRODUÇÃO
As principais características dos Moneras são:unicelulares; procariontes (isto é, possuem célula procariótica, desprovida de núcleo organizado e organelas membranosas);
· metabolismo autótrofo (fotossintetizantes ou quimiossintetizantes) ou heterótrofo;
· aeróbicos ou anaeróbicos.
Os seres do Reino Monera são sempre unicelulares, embora em muitos casos se mostrem como coloniais, formando aglomerados celulares. Nas cianobactérias particularmente, esses aglomerados são, por vezes, tão bem organizados que aparentam ser organismos pluricelulares filamentosos.
Comece a conhecer o Reino Monera
assistindo ao vídeo sobre as bactérias:
"Tudo sobre Bactérias - Programa Discovery na Escola (Discovery Channel)
2. INTEGRANTES: ARCHAEA e BACTERIA (eubactérias)
Separação dos moneras em dois grupos, Bacteria e Archaea (geralmente anaeróbicos quimiossintetizantes), ocorreu na década de 1970, graças as descobertas do microbiologista Carl Woese, utilizando comparação genética. Por meio de estudos comparativos de sequências de DNA que codificavam RNAr de diferentes seres, foram definidos 3 grandes domínios incluindo todos os seres do Planeta. Assim, de acordo com a proposta de Woese, os seres poderiam ser agrupados em três grandes domínios: Bacteria (anteriormente denominados eubactérias), Archaea (ou arqueas, anteriormente denominadas “arqueobactérias”) e Eucarya (eucariontes). Estes três domínios teriam derivado de um hipotético ancestral comum de todas as células.
REINO MONERA
|
|
ARCHAEA
|
BACTERIA
|
Termofílicas
Halofílicas
Metanogênicas
|
bactérias
cianobactérias |
3. ARCHAEA
Ao contrário das bactérias, as Archeas não têm parede celular de peptidoglicano e o flagelo de arquea tem composição química diferente.
As proteínas de membrana podem ser também bastante diferentes daquelas observadas em outros tipos celulares. Veja algumas diferenças:
REINO MONERA
| |
ARCHAEA
|
BACTERIA
|
de outros seres (vivos ou em decomposição) Pequeno número de espécies habita ambientes de condições extremas. - Membrana composta por fosfolipídeos e colesterol. - Ribossomos 30S e 60S.; - RNA polimerase - de 4 tipos. | |
A maioria é encontrada em poucos ambientes terrestres e aquáticos pouco favoráveis, tais como lagos extremamente salinos, ambientes estritamente anaeróbios e/ou ambientes extremamente quentes. O conhecimento sobre este grupo vem mostrando que as características apresentadas por alguns Archaea parecem refletir as condições da Terra primitiva, quando tal domínio começou a evoluir como um ramo filogenético distinto. Ao que parece, várias arqueas conservaram, mais do que as eubactérias, estas características fisiológicas primitivas, o que seria responsável pela distribuição atual no Planeta. Assim, as Archaea compreendem um grupo heterogêneo de microrganismos que podem ser caracterizados, em sua maioria, como habitantes de ambientes inóspitos, geralmente crescendo em condições consideradas até então como extremas e limítrofes para a vida. De modo geral, e pelo fato da maioria das Archaea conhecidas serem isoladas de ambientes quentes, salinos e/ou anaeróbios, este domínio é didaticamente dividido em três grupos: termofílicos, halófilos e metanogênicos. No entanto, vale relembrar mais uma vez que dados mais recentes mostram ampla distribuição geográfica e ecológica desse grupo de organismos. Archaea podem ser encontradas nos mais diversos ecossistemas, desde ambientes aquáticos frios até no sistema digestório de animais, incluindo o homem e tecidos vegetais.
Atualmente, sabemos que estes seres são importantes membros da microbiota aquática de regiões frias do planeta. Ao que parece, as Archaea podem corresponder a até 34% da biomassa procariótica das águas costeiras superficiais da Antártida.
Termofílicas: Suportam viver em ambientes com temperatura elevada, Pyrolobus fumarii foi isolada de uma fenda no Oceano Atlântico, a uma profundidade de
As termófilas podem ser encontradas em fontes geotérmicas, fontes vulcânicas (que expelem vapores e compostos sulfurados), fontes termais marinhas, onde erupções vulcânicas elevam a temperatura para mais de

Fonte termal marinha. Disponível em: http://www.spaceref.com/news/viewpr.html?pid=6101

A quantidade de sal no Mar Morto é tal que é possível se manter facilmente na superfície. Comparando-o com a concentração média de outros oceanos em que o teor de sal, por 100 ml de água, não passa de 3 g, no
Mar Morto fica entre 30 a 35 g de sal/100 ml de água, ou seja, dez vezes
superior.

Acúmulo de sal
4. BACTERIA
Esse é o maior grupo de moneras e inclui bactérias e cianobactérias.
4.1. BACTÉRIAS
A bioquímica variada das diferentes espécies de bactérias está relacionada à sua sobrevivência em quase todos os ambientes, muitas vezes em circunstâncias onde somente moléculas muito simples estão disponíveis para o metabolismo de base. Constituem os seres mais disseminados pela face da Terra, vivendo nos mais variados ambientes – aéreo, terrestre, aquático.
TEMPERATURA
|
INFLUÊNCIA NA MANUTENÇÃO DEBACTÉRIAS
|
Abaixo de 6ºC
|
As bactérias não se multiplicam, mas não morrem.
|
Entre 7ºC e 16ºC
|
As bactérias multiplicam-se lentamente.
|
Entre 17ºC e 50ºC
|
As bactérias multiplicam-se rapidamente.
|
Entre 51ºC e 80ºC
|
Acima de 100ºC
|
Praticamente todas as bactérias morrem.
|
4.1.1 MODO DE VIDA

* Saprófitas: vivem às custas da matéria orgânica livre no ambiente, em decomposição. Ex.: bactérias que decompõem a serrapilheira na Mata Atlântica.
* Comensais: vivem às custas da matéria orgânica viva, associadas a seres dos quais retiram nutrientes, porém não há prejuízo nem benefício àquele ser que hospeda esta categoria de bactérias. Ex.: bactérias da microbiota* intestinal.
* Parasitas: vivem às custas da matéria orgânica viva, associados a seres dos quais retiram nutrientes, prejudicando-os. Ex.: bactérias causadoras de doenças.
* Simbiontes: vivem associadas a outros seres, numa espécie de "contrato com benefícios mútuos". Ex.: bactérias da microbiota intestinal humana, bacterioriza.
* microbiota intestinal: conjunto de microrganismos que colonizam a luz intestinal; algumas são comensais, outras realizam tarefas que acabam trazendo benefícios ao homem, tal como a síntese de vitaminas e de aminoácidos essenciais. O termo substitui o antigo "flora intestinal", uma vez que "flora" designa conjunto de plantas. O termo "flora" é do tempo em que as bactérias eram estudadas pelos botânicos.
4.1.2 TAMANHO
As bactérias são visíveis somente ao microscópio ótico, utilizando-se aumento médio de 1.000 vezes, pois a maioria das bactérias mede cerca de 1Mm (micrômetro) de diâmetro. É difícil compreender, realisticamente, esse tamanho. Podemos comparar com uma semente de feijão que, se aumentada 1000 vezes, ficaria com o diâmetro de 10 metros.
1 Mm = 10-3 mm
As menores e mais simples bactérias conhecidas são as riquétsias, clamídias e os micoplasmas.
As riquétsias são seres tão inferiores que são consideradas por muitos biólogos como um meio-termo entre vírus e bactérias; dificilmente visíveis ao microscópio comum; como os vírus. são parasitas intracelulares. Cerca de 50 espécies de riquétsias são parasitas inócuos no trato intestinal e glândulas salivares de insetos tais como piolhos, barbeiros e carrapatos.O tifo epidêmico é causado por Rickettsia prowazekii, transmitida ao homem por piolhos – Pediculus humanus e chatos (piolho pubiano) contaminados com essas bactérias.
As clamídias também são parasitas intracelulares, são transmitidas aos humanos de forma direta, de pessoa para pessoa. São causadoras do tracoma (infecção nos olhos, muito contagiosa, caracterizada por inflamação das conjuntivas e da córnea, causada pela bactéria Chlamydia trachomatis) e do linfagranuloma venéreo.
Os micoplasmas são menores ainda que as
riquétsias, em média, do tamanho dos maiores vírus; são as menores células conhecidas. Não têm parede celular, o que torna a célula frágil e deformável, mas é também a razão para o fato de a penicilina ter pouco efeito sobre elas.

As riquétsias são seres tão inferiores que são consideradas por muitos biólogos como um meio-termo entre vírus e bactérias; dificilmente visíveis ao microscópio comum; como os vírus. são parasitas intracelulares. Cerca de 50 espécies de riquétsias são parasitas inócuos no trato intestinal e glândulas salivares de insetos tais como piolhos, barbeiros e carrapatos.O tifo epidêmico é causado por Rickettsia prowazekii, transmitida ao homem por piolhos – Pediculus humanus e chatos (piolho pubiano) contaminados com essas bactérias.
As clamídias também são parasitas intracelulares, são transmitidas aos humanos de forma direta, de pessoa para pessoa. São causadoras do tracoma (infecção nos olhos, muito contagiosa, caracterizada por inflamação das conjuntivas e da córnea, causada pela bactéria Chlamydia trachomatis) e do linfagranuloma venéreo.
 |
| Tracoma |

A bactéria simbionte do
intestino do peixe-cirurgião Acanthurus chirurgus mede cerca 0,5 milímetro
de comprimento e é visível a olho nu.
4.1.3 FORMA
Embora existam
milhares de espécies bacterianas diferentes, as bactérias isoladas têm uma das
4 formas básicas: - esférica (cocos); - bastonete (bacilo); - espiralada
(espirilo); - forma de vírgula (vibrião).
| Forma das bactérias livres e coloniais. |
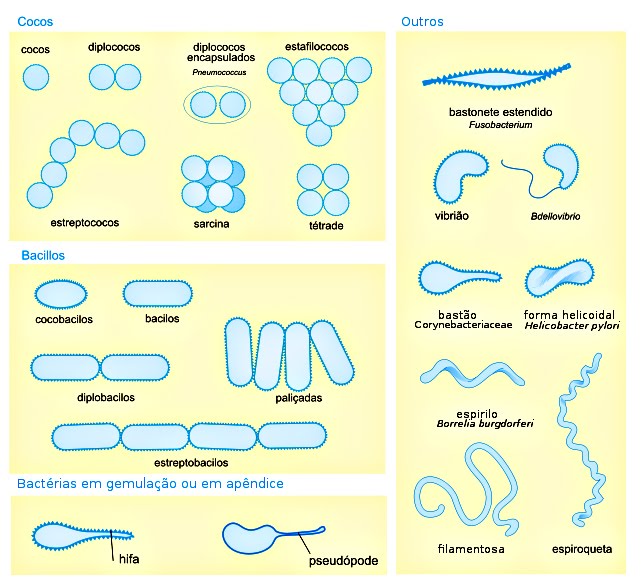 |
4.1.4 ESTRUTURA
Parede Celular
As bactérias, com exceção de alguns grupos (uma exceção é o micoplasma), apresentam parede celular espessa e rígida envolvendo, dando forma à célula e protegendo contra choques mecânicos e osmóticos (em meio hipotônico). Essa parede é permeável, permitindo a troca de substâncias entre a célula e o meio externo. Sua constituição química é diferente da encontrada nos fungos (principalmente quitina) e nas plantas (celulose) e varia nas diferentes espécies de bactérias.
As bactérias, com exceção de alguns grupos (uma exceção é o micoplasma), apresentam parede celular espessa e rígida envolvendo, dando forma à célula e protegendo contra choques mecânicos e osmóticos (em meio hipotônico). Essa parede é permeável, permitindo a troca de substâncias entre a célula e o meio externo. Sua constituição química é diferente da encontrada nos fungos (principalmente quitina) e nas plantas (celulose) e varia nas diferentes espécies de bactérias.
Em
1884, o bioquímico dinamarquês Hans Christian Gram descobriu que bactérias sem
uma camada de lipídios associada a polissacarídeos na parede celular absorviam
o corante violeta de genciana. Tal processo, chamado coloração Gram, é usado para classificar as bactérias em
Gram-positivas ou Gram-negativas. Estudos mostraram que certos antibióticos são
mais eficientes contra
bactérias Gram +, enquanto outros são indicados para tratamento de infecções
causadas por bactérias Gram negativas.
A parede celular das bactérias pode ser formada primariamente por uma espessa camada de peptidoglicano (moléculas de açúcar associadas a aminoácidos). As bactérias com esse tipo de parede celular, são denominadas gram-positivas (Gram +), pois retêm o corante violeta utilizado para diferenciar bactérias (método de Gram). As bactérias Gram+ são menos resistentes.
Em outras bactérias, a parede celular é formada por uma camada delgada de peptidoglicano e uma camada externa adicional, semelhante a uma segunda membrana plasmática formada por lipídios unidos a polissacarídeos, formando lipopolissacarídeo. Essas bactérias são denominadas gram-negativas, pois não retêm o referido corante. Essas bactérias são mais resistentes à penicilina e outros antibióticos. O antibiótico penicilina provoca a morte da célula bacteriana, pois interfere na síntese do peptidoglicano, e as bactérias não conseguem sobreviver sem a parede celular para protegê-las. A penicilina é mais efetiva contra as bactérias
gram-positivas.
Membrana plasmática
Cápsula
A parede celular das bactérias pode ser formada primariamente por uma espessa camada de peptidoglicano (moléculas de açúcar associadas a aminoácidos). As bactérias com esse tipo de parede celular, são denominadas gram-positivas (Gram +), pois retêm o corante violeta utilizado para diferenciar bactérias (método de Gram). As bactérias Gram+ são menos resistentes.
Em outras bactérias, a parede celular é formada por uma camada delgada de peptidoglicano e uma camada externa adicional, semelhante a uma segunda membrana plasmática formada por lipídios unidos a polissacarídeos, formando lipopolissacarídeo. Essas bactérias são denominadas gram-negativas, pois não retêm o referido corante. Essas bactérias são mais resistentes à penicilina e outros antibióticos. O antibiótico penicilina provoca a morte da célula bacteriana, pois interfere na síntese do peptidoglicano, e as bactérias não conseguem sobreviver sem a parede celular para protegê-las. A penicilina é mais efetiva contra as bactérias
gram-positivas.
Membrana plasmática
Sob a parede
celular, encontra-se a membrana plasmática, lipoprotéica, com estrutura espacial na forma de "mosaico fluido", a mesma das demais
células. Na face interna da membrana plasmática encontram-se
várias enzimas relacionadas com o metabolismo bacteriano.
Citoplasma
O citoplasma das bactérias é formado por
água e diversas substâncias inorgânicas e orgânicas; não apresenta
microtúbulos e microfilamentos formando um citoesqueleto, como ocorre no
citoplasma das células eucarióticas. Dentre as organelas
citoplasmáticas presentes nas células eucarióticas, apenas os ribossomos
ocorrem também nas células procarióticas; as organelas membranosas não estão
presentes. A única estrutura membranosa presente no citoplasma da maioria das
bactérias é o mesossomo, que
corresponde simplesmente a uma invaginação da membrana plasmática. Os
mesossomos aumentam a superfície da membrana e atuam como locais de
concentração de enzimas, principalmente daquelas relacionadas ao processo de
respiração; além disso, o DNA da bactéria está, em geral, ligado ao mesossomo.
Imerso no citoplasma,
encontra-se o material genético (cromossomo), formando uma estrutura
denominada nucleoide. O cromossomo é
uma molécula circular de DNA, não associada a proteínas. Nas células
eucarióticas, o cromossomo não é circular, e o DNA associa-se a proteínas. Além disso, os cromossomos das
células eucarióticas encontram-se no interior do núcleo, separados do
citoplasma pela carioteca. A carioteca é ausente em todos
os procariontes.
Além do nucleoide (DNA
principal), podem ocorrer no citoplasma das células bacterianas, moléculas
menores e circulares de DNA - os plasmídeos
(plasmídio = pedaço de DNA
circular que está dentro do nucleóide; epissoma = pedaço de DNA que está fora
do nucleóide) O plasmídio é autonômo, duplica-se
independentemente do DNA do nucleóide.
Dependendo do tipo de
bactéria, pode haver um ou muitos plasmídeos por célula. Os plasmídeos não têm os genes essenciais para o
funcionamento da célula, mas podem apresentar genes que comandam a síntese de
proteínas capazes de degradar as moléculas dos antibióticos, conferindo
resistência às bactérias.
Cápsula
Nas células bacterianas pode
haver, externamente à parede celular, uma cápsula
formada por substâncias viscosas, produzidas pela própria célula. A cápsula não
é um componente obrigatório. Essa cápsula
atua como envoltório protetor, dificultando a fagocitose por parte dos
glóbulos brancos, aumentando o poder infectante nas espécies patogênicas.
Nestas, a perda da cápsula pode diminuir ou mesmo eliminar seu poder
infectante. Além de conferir uma proteção extra à bactéria contra a penetração
de vírus e contra o ataque de glóbulos brancos, a cápsula facilita a adesão da
bactéria em vários tipos de superfícies – inclusive nos dentes, no caso das
bactérias que provocam a cárie.
A maior parte das cápsulas é formada por
polissacarídeos, embora existam cápsulas formadas por polipeptídeos ou outros
compostos.
A constituição química da cápsula é altamente específica. Por exemplo, existem cerca de 70 tipos diferentes de pneumococos, identificados com base em pequenas alterações na composição química das cápsulas.
A constituição química da cápsula é altamente específica. Por exemplo, existem cerca de 70 tipos diferentes de pneumococos, identificados com base em pequenas alterações na composição química das cápsulas.

Bacilo - desenho de Eliz Garcia Ferreira (2013)
As bactérias podem ou não apresentar movimentos; as que se movimentam o fazem através de flagelos ou por deslizamento. Os flagelos ocorrem principalmente nos bacilos e nos espirilos, podendo haver um ou mais flagelos por célula.
Fímbria
Algumas bactérias podem apresentar fímbrias (ou Pili), estruturas menores e mais
numerosas que os flagelos e que não
desempenham nenhum papel relativo à mobilidade, podendo
ocorrer tanto em bactérias móveis como em imóveis. As funções
das fímbrias parecem estar associadas à adesão das células bacterianas às
diferentes superfícies de contato. Há uma fímbria especial – a fímbria
sexual, cuja função é a de formar uma ponte citoplasmática entre bactérias
em processo de conjugação (troca de material genético entre bactérias).
4.1.5 NUTRIÇÃO
A maioria das bactérias é heterótrofa por absorção, retirando moléculas orgânicas do ambiente (saprófitas) ou de seres vivos que parasitam (parasitas), ou ainda a partir de uma relação de simbiose (por exemplo, nas raízes das leguminosas).
Algumas são autótrofas por fotossíntese (usam a energia luminosa para sintetizar compostos orgânicos) ou por quimiossíntese (usam energia de reações químicas para produzir compostos orgânicos – nitro, sulfo e ferrobactérias). As bactérias fotossintetizantes possuem um pigmento chamado bacterioclorofila, quimicamente semelhante à clorofila das células vegetais. As cianobactérias possuem clorofila idêntica à encontrada nas plantas. A fotossíntese bacteriana é diferente da realizada pelas plantas. Em primeiro lugar, não há consumo de H2O como fonte de H para a síntese de glicose; em segundo, não libera O2.
Fotossíntese das plantas:
6CO2 + 12H2O __energia luminosa ® C6H12O6 + 6H2O + 6O2
Fotossíntese de bactérias:
6CO2 + 12H2S __energia luminosa ® C6H12O6
+ 6H2O + 12S6CO2 + 12H2O __energia luminosa ® C6H12O6 + 6H2O + 6O2
Fotossíntese de bactérias:
Observações: a fotossíntese dessas bactérias utiliza gás sulfídrico como fonte de H para a síntese de glicose; utiliza o pigmento bacterioclorofila para a captura da energia luminosa e não libera oxigênio.
As bactérias quimiossintetizantes utilizam a energia de reações químicas, em vez de energia luminosa, para a síntese de matéria orgânica a partir da matéria inorgânica. Elas utilizam energia obtida pelas reações de oxidação de compostos inorgânicos, que realizam previamente.
Nitrobacter, sulfobactérias e
ferrobactérias são quimiossintetizantes
e vivem em fontes de águas termais ou no solo, onde encontram substratos
minerais para as suas reações iniciais de oxidação.
Nitrobacter e Nitrosomonas participam da reciclagem do N no
planeta.

Disponível em: http://pt.wikipedia.org/wiki/Ficheiro:Nitrogen_Cycle_pt.png
Nitrobactérias: oxidam nitrito em nitrato, são bactérias que vivem no solo, o NO3
formado é absorvido pelas raízes das plantas. Esse NO3 é reduzido no
interior da planta e transformado em matéria orgânica nitrogenada, como aminoácidos e proteínas.
NITRATAÇÃO
2NO-2
+ O2 ® 2NO3-
+ 2H+ + ATP
nitrito nitrato
Nitrosomonas e
Nitrosococcus: oxidam amônia (gasoso; ou amônio - NH4 dissolvido em água) em nitrito:
NITROSAÇÃO
2NH3 + 3O2 ® 2NO2 + 2H+
+ 2H2O + ATP
amônia (gasoso) nitrito

Sulfobactérias: oxidam o gás sulfídrico em duas etapas:
2H2S + O2 ® 2H2O +2S + ATP
Numa segunda etapa, elas oxidam o S
liberado na primeira reação e que está acumulado no citoplasma, produzindo
ácido sulfúrico e ATP.
2S + 2H2O
+ 3O2 ® 2H2SO4
+ ATP
Depois utilizam o ATP para construir glicose:
6CO2 + 6H2O ® C6H12O6 + 6O2
Ferrobactérias: 2FeCO3 + 3H2O + ½ O2 ® 2Fe(OH)3 + 2CO2 + ATP
4.1.6 RESPIRAÇÃO
A respiração celular é o
processo através do qual os seres vivos conseguem energia para realizar suas
atividades, oxidando compostos orgânicos – a glicose. A maioria dos seres usa o O2
para extrair energia da glicose – é a respiração
aeróbica (do grego ero = ar).
Outros seres, porém, podem
conseguir energia sem usar O2 – através da respiração anaeróbica.
Esse processo é usado, por exemplo, por bactérias que vivem no solo ou em águas
estagnadas, onde o suprimento de oxigênio é escasso. Os produtos finais da
respiração anaeróbica são o CO2 e uma substância usada como reagente
em lugar do O2. Por exemplo, certas bactérias do solo, chamadas
desnitrificantes, como Pseudomonas, usam nitratos na respiração anaeróbica
e liberam nitrogênio molecular (N2) para o ar. À transformação dos
nitratos em N2 dá-se o nome de desnitrificação.
Algumas bactérias também podem obter
ATP sem utilizar de O2, através de outros processos anaeróbicos como a fermentação (é o metabolismo anaeróbico de carboidratos chegando-se a ácido acético: fermentação acética, Acetobacter (vinagre) oxida álcool etílico em ácido acético e a ácido láctico: fermentação láctica, Lactobacyllus - iogurte) e a putrefação (é o metabolismo anaeróbico de obtenção de ATP a partir de proteínas e lipídios) que não podem ser considerados como respiração anaeróbica. A palavra respiração
é empregada em todo processo de síntese de ATP que envolve a cadeia
respiratória.
As
bactérias podem ser divididas, quanto à respiração, em 3 grupos:
aeróbicas: dependem do O2
para conseguir energia:
Respiração aeróbica
C6H12O6
+ 6 O2 + 38 ADP + 38 Pi ® 6 CO2+ 6 H2O + 38 ATP
anaeróbicas
Anaeróbicas facultativas: vivem com ou sem O2; se houver O2 no ambiente, podem realizar respiração aeróbica; caso contrário, sobrevivem à custa de processos anaeróbicos. As bactérias anaeróbicas também podem obter energia através da fermentação e da putrefação. Na fermentação, a obtenção de energia é feita pela degradação parcial de carboidratos, chegando-se a ácido acético (fermentação acética), a ácido láctico (fermentação láctica) ou a álcool etílico (fermentação alcoólica). A putrefação é o processo anaeróbico que degrada proteínas ou lipídios. A quantidade de energia obtida é sempre inferior à da respiração aeróbica.
Anaeróbicas facultativas: vivem com ou sem O2; se houver O2 no ambiente, podem realizar respiração aeróbica; caso contrário, sobrevivem à custa de processos anaeróbicos. As bactérias anaeróbicas também podem obter energia através da fermentação e da putrefação. Na fermentação, a obtenção de energia é feita pela degradação parcial de carboidratos, chegando-se a ácido acético (fermentação acética), a ácido láctico (fermentação láctica) ou a álcool etílico (fermentação alcoólica). A putrefação é o processo anaeróbico que degrada proteínas ou lipídios. A quantidade de energia obtida é sempre inferior à da respiração aeróbica.
Fermentação láctica
C6H12O6
+ 2 ADP + 2Pi ® 2 C3H6O3
(ácido láctico) + 2 ATP
Fermentação alcoólica
C6H12O6
+ 2 ADP + 2Pi ® 2 C2H5OH
(álcool etílico) + 2 CO2+ 2 ATP
Anaeróbicas obrigatórias (restritas): não têm enzimas para o
aproveitamento de O2 e, por isso, morrem a partir de determinada
concentração de O2, tais como algumas
bactérias do solo, algumas bactérias do fundo do mar e algumas bactérias
patogênicas (causadoras de doenças).
Ex.:
Clostridium tetanii é anaeróbica restrita. O hábito de
colocar água oxigenada (H²O²) sobre ferimentos faz
liberar O2 pela ação da catalase (uma peroxidade) liberada dos peroxissomos
das células lesadas –2 H²O² + catalase = O² +2 H²O.
A respiração anaeróbica
envolve um receptor de elétrons diferente do oxigênio e existem vários tipos de
bactérias capazes de usar uma grande
variedade de compostos como aceptoras
na respiração: compostos nitrogenados, tais como nitratos e nitritos,
compostos de enxofre, tais como sulfatos,
sulfitos, dióxido de enxofre e mesmo
enxofre elementar, dióxido de carbono, compostos de ferro, de manganês, de
cobalto e até de urânio. No entanto, para todos
estes, a respiração anaeróbica só ocorre em ambientes onde o oxigênio é
escasso, como nos sedimentos marinhos, lacustres ou naqueles próximos de nascentes
hidrotermais submarinas.
Veja o caso das bactérias do gênero Desulfovibrio, que utilizam sulfato como
composto aceptor:
Respiração anaeróbica em Desulfovibrio desulfurican
2C3H5O3- + SO42- Æ 2C2H3O2- + 2CO2 + HS- + 2ATP
lactato sulfato acetato
Respiração
anaeróbica de bactéria desnitrificante
C6H12O6 + 4 NO3- Æ 6CO2 + 6H2O + 2N2 +
energia
nitrato
ASSEXUADA
As bactérias
reproduzem-se por um mecanismo assexuado em que uma bactéria dá
origem a outras duas, geneticamente idênticas. Esse tipo de reprodução
assexuada pode ser denominado bipartição
ou cissiparidade, embora não seja
semelhante à mitose dos eucariontes, pois não há condensação dos cromossomos,
não há formação de fuso mitótico (os microtúbulos são ausentes nos procariontes)
e não há carioteca. Na divisão
da célula bacteriana, o cromossomo duplica-se e se forma mais um mesossomo.
Cada cromossomo fica associado a um mesossomo e entre eles verifica-se o
crescimento da célula. A seguir, ocorre a citocinese (divisão do citoplasma), dando origem a duas
células-filhas.




A transdução ocorre quando um vírus, bacteriófago, ao se reproduzir por lisogenia dentro de bactérias, pode sair contaminado por pedaços de DNA bacteriano, transportando-as para outra bactéria.
As bactérias têm alto poder
de reprodução. Em algumas horas, sob condições ambientais adequadas, uma única
célula pode dar origem a milhares de descendentes, todos geneticamente
idênticos entre si.
"SEXUADA"
Alterações genéticas nas
bactérias ocorrem por mutação e, em alguns casos, por transmissão de material
genético durante a recombinação genética, podendo ocorrer através de três
mecanismos distintos:
- conjugação;
- transformação e
- transdução.
Destes, a
conjugação bacteriana é o mecanismo mais frequentemente utilizado pelas bactérias. A conjugação bacteriana foi
descoberta quando duas variedades geneticamente diferentes da bactéria Escherichia
coli foram cultivadas juntas em laboratório. Os pesquisadores verificaram que,
com alguma freqüência, células de uma variedade uniam-se temporariamente com
células da outra variedade, através de uma ponte citoplasmática. Analisando o processo com
mais detalhes, pode-se constatar que em uma das células, denominada doadora ou macho(é
dito F+),, ocorre duplicação de parte de seu cromossomo. Essa porção duplicada (o
fator F) separa-se do cromossomo original e passa, através da ponte
citoplasmática, para a outra célula, que é denominada receptora ou fêmea (não
contém o fator F = fertilidade, é dita F-). Essa porção do cromossomo une-se ao
cromossomo da célula receptora, que fica, então, com constituição genética
diferente. Essa bactéria “recombinante” pode dividir-se assexuadamente várias
vezes, formando um clone geneticamente distinto dos produzidos pelas bactérias
“não-recombinantes”.
A conjugação é considerada
uma forma de reprodução sexuada, uma vez que ocorre recombinação de genes entre
indivíduos, o que aumenta a variabilidade genética de uma espécie.

Na transformação uma bactéria pode
absorver DNA livre do meio ambiente (proveniente de outra bactéria morta) e
incorporá-lo ao seu material genético.

A transdução ocorre quando um vírus, bacteriófago, ao se reproduzir por lisogenia dentro de bactérias, pode sair contaminado por pedaços de DNA bacteriano, transportando-as para outra bactéria.

4.1.8 FORMAÇÃO de ESPOROS
Algumas bactérias formam, em condições adversas, estruturas resistentes chamadas esporos, que suportam condições críticas de temperatura e falta de água que normalmente levariam à morte. Essas estruturas são células de parede resistente, com pouca água no citoplasma, metabolismo reduzido; nesse caso a bactéria fica latente, com as funções vitais reduzidas ao mínimo. Quando as condições do meio voltam à normalidade, a bactéria retoma suas atividades biológicas.
4.1.9 IMPORTÂNCIA das BACTÉRIAS
Nos ecossistemas
A maior parte das bactérias
obtém seus alimentos degradando matéria orgânica morta. Essas bactérias são
denominadas decompositoras ou saprófitas e, juntamente com os fungos,
são responsáveis pela decomposição e reciclagem da matéria orgânica nos
diferentes ambientes sendo, assim, importantíssimas para o equilíbrio
ecológico de ecossistemas marinhos (talassociclo), dulcícolas (limnociclo) e terrestres (epinociclo).

Algumas digerem a celulose no tubo digestivo dos ruminantes.
As bactérias
também têm importância ecológica no chamado ciclo do nitrogênio. O
nitrogênio atmosférico, antes de fazer parte de aminoácidos, proteínas e ácidos
nucleicos, precisa ser transformado em substâncias aproveitáveis pelas plantas.
As
bactérias do gênero Rhizobium têm notável capacidade de reter o nitrogênio
livre da atmosfera, formando nitratos. Em virtude disso, elas se associam
rotineiramente com plantas leguminosas (feijão, soja, ervilha, alfafa, etc),
formando nódulos nas raízes dessas plantas – as conhecidas bacteriorrizas.
Em cada nódulo, há muitos milhares de bactérias, as quais se nutrem dos hidratos
de carbono produzidos pelas leguminosas no processo de fotossíntese, mas
oferecem, em troca, os nitratos, que a planta não tem capacidade de produzir.
Os nitratos constituem fonte de nitrogênio para que a planta construa seus
aminoácidos e proteínas. É um caso de mutualismo. É fato comprovado que
as leguminosas têm crescimento precário em solos pobres de bactérias do gênero Rhizobium.


A B C
A - nódulos de Rhizobium nas raízes; B -planta em solo com bactérais Rhizobium; C - plantas em solo previamente tratado com antibióticos.
Os compostos nitrogenados
produzidos pelas bactérias dos nódulos acabam fertilizando o solo, o que
favorece também as plantas não-leguminosas. Por isso certos agricultores
alternam plantio de espécies não-leguminosas, como o milho, por exemplo, com o
plantio de leguminosas, método conhecido como rotação de cultura.
Pode-se plantar leguminosas e não-leguminosas ao mesmo tempo, em fileiras
alternadas, método conhecido como plantação consorciada.
4.1.20. DOENÇAS por BACTÉRIASAlgumas espécies de bactérias são causadoras de doenças em vários grupos de seres vivos, entre eles o grupo das plantas e dos animais, como o homem.
Para facilitar a memorização da DOENÇAS BACTERIANAS veja a dica de Renato Almeida no Canal Digital Academy:
- Habitat: maioria dulcícola (Anabaena cheiro e gosto desagradável à
água), solo úmido, água salgada (mares), salobra (manguezais), pelos de animais
(urso polar, preguiça), troncos. Algumas espécies ocorrem em fontes
termais (80ºC), suportam também seca intensa e excesso de radiação solar.
- Modo de vida:
autótrofas fotossintetizantes; fotossíntese semelhante à de plantas e algas
protistas. Simbiontes – liquens.
- Estrutura: célula procariótica; células
isoladas, outras formam colônias filamentosas com bainha
mucilaginosa externa à parede celular; vacúolos gasosos
envolvidos por camada proteica, e não membrana lipoprotéica, permitem flutuação
na superfície para captura de luz. Ausência de flagelos e fímbrias.
O aparelho fotossintético mais organizado do que o de bactérias
fotossintetizantes. Pigmentos ficam em lamelas, dobras da membrana,
sobre as quais há grânulos chamados cianossomas ou ficobilissomas (ficocianina –azul;
ficoeritrina -vermelho). A
cianobactéria utiliza, de maneira eficiente, um espectro de luz mais amplo, que
não seria possível apenas com a clorofila.

http://www.vestibulandoweb.com.br/biologia/teoria/celula.asp
Os pigmentos ficobilinas, assim como a capacidade de formar esporos
resistentes, facilita o crescimento sob condições que impediriam a
sobrevivência da maioria dos vegetais superiores.
- Reprodução: assexuada, por bipartição; formas
filamentosas podem reproduzir-se assexuadamente por fragmentação: quebram-se em alguns pontos, dando origem a vários
fragmentos pequenos chamados hormogônios,
que, por divisão de suas células, darão origem a novas colônias filamentosas.
Formas coloniais produzem esporos resistentes - os acinetos (armazena
substâncias de reserva)., que podem destacar-se e originar novos filamentos.


Disponível em: http://mundociano.blogspot.com.br/2011/12/cyanobacteria.html
- Fixadoras de N: assim como as bactérias, têm importância ecológica, pois também fixam
o N2 do ar para a síntese de compostos nitrogenados
indispensáveis à sua nutrição. O fato de
poderem fixar N2 do ar e realizar a fotossíntese permite a
sobrevivência das cianofíceas em ambientes muito simples, onde haja apenas N2,
CO2, água, luz e poucos sais minerais.
Constituem, por isso, espécies pioneiras, capazes de implantar a vida em
regiões estéreis. Uma célula especial – o heterocisto,
que ocorre intermitentemente ao longo da cadeia de células do filamento, cuja
função é fixar nitrogênio e distribuir as outras células do filamento.
Os ficobilissomos não são encontrados nos heterocistos.
Algumas espécies liberam toxinas na água que
nem os meios mais eficazes de descontaminação conseguem limpar a água. Essas
toxinas podem ser do tipo hepatotoxina ou neurotoxina.
Nenhum comentário:
Postar um comentário